Abstract
Background. Pseudomonas aeruginosa biofilm polymer matrix formation contributes to antibiotic tolerance. The antibiofilm effects of sub-minimum inhibitory concentrations (MICs) of ceftriaxone (CTX), the molecular mechanisms by which these sub-MICs modulate biofilm polymer production and quorum sensing (QS), and the binding interactions of CTX with key biofilm regulatory proteins (LasR and RhlR QS receptors) have not been previously investigated.
Objectives. To determine the role of sub-MIC CTX in regulating biofilm polymer matrix formation, bacterial adhesion, QS gene expression (rhlR and lasR), and to perform molecular docking analysis of CTX interactions with LasR and RhlR QS receptor proteins and biofilm EPS polymer-associated targets.
Materials and methods. MICs and biofilm formation were determined. The effects of CTX sub-MICs on biofilm formation, adhesion to mouse bladder epithelial cells (BECs), and QS gene expression (rhlR and lasR, by qRT-PCR) were assessed. In silico molecular docking of CTX against the ligand-binding domains of LasR (PDB: 2UV0) and RhlR (PDB: 3T5K) was performed using AutoDock Vina. Interaction fingerprinting with biofilm EPS polymer-associated enzymes (AlgD and PelB) was also performed.
Results. CTX sub-MICs regulated biofilm formation in an isolate-dependent manner, reduced P. aeruginosa adhesion to mouse BECs, and downregulated the rhlR and lasR genes in a concentration-dependent manner. Molecular docking revealed that CTX binds favorably within the ligand-binding pockets of LasR (−8.3 kcal/mol) and RhlR (−7.1 kcal/mol) via hydrogen bonding and hydrophobic interactions, suggesting competitive interference with QS autoinducer binding. CTX also exhibited affinity for AlgD (−7.6 kcal/mol), a key enzyme in alginate polymer biosynthesis.
Conclusions. CTX sub-MICs modulate biofilm EPS polymer matrix formation and epithelial adhesion by downregulating QS regulatory genes. lasR was more responsive to CTX sub-MIC stress than rhlR. Molecular docking supports a direct molecular interaction mechanism through which CTX may interfere with QS receptor signaling and alginate polymer biosynthesis, providing a structural basis for its antibiofilm activity at sub-inhibitory concentrations.
Key words: Pseudomonas aeruginosa, alginate, molecular docking, ceftriaxone, quorum sensing
Background
The formidable opportunistic pathogen Pseudomonas aeruginosa causes persistent infections in immunosuppressed patients and in individuals with cystic fibrosis, burns, urinary tract infections (UTIs), and infections associated with indwelling medical devices.1, 2 Its primary virulence factor is its ability to form biofilms. A biofilm is a structured, multicellular community of bacteria enclosed within a self-produced extracellular matrix. This mode of growth creates a microenvironment that promotes bacterial proliferation and shields the bacteria from antibiotics and the host immune system.3 Consequently, it often leads to chronic and difficult-to-treat infections.4 This inherent resistance to antibiotics contributes to high morbidity and mortality rates, presenting a significant challenge for managing P. aeruginosa infections in clinical settings.5
The extracellular matrix of P. aeruginosa biofilms is a complex mixture of biopolymers that confer structural stability, antibiotic recalcitrance, and protection from host immunity. Chief among these are three distinct exopolysaccharide (EPS) polymers: alginate, Pel, and Psl. 6 Alginate is a linear polymer of β-1,4-linked mannuronate and guluronate residues, which forms a viscous, hydrogel-like capsule that impedes antibiotic penetration and protects bacteria from desiccation and immune surveillance.7 Pel is a cationic, partially deacetylated polymer of β-1,4-linked galactosamine and N-acetylgalactosamine, critical for maintaining biofilm cohesion and pellicle formation. Psl is a neutral pentasaccharide repeat-unit polymer rich in mannose and glucose, predominantly associated with surface attachment and the initial stages of biofilm development.8 Together, these EPS polymers represent the structural ‘scaffold’ of the biofilm and are prime targets for anti-biofilm therapeutic strategies. Understanding how antibiotics at sub-inhibitory concentrations alter EPS polymer production is therefore critical for comprehending biofilm modulation.
The cell-to-cell communication system known as quorum sensing (QS) is involved in the pathogenicity of P. aeruginosa. The Las and Rhl systems are the primary circuits in P. aeruginosa that function hierarchically yet interactively. Both systems have a transcriptional regulator (LasR and RhlR, respectively) and its cognate autoinducer synthase.9 The LasR system is the primary regulator that drives activation of the RhlR system. Both systems regulate the expression of various virulence factors, including elastase, rhamnolipids, and pyocyanin, and are essential for the formation and maintenance of strong biofilms.10 Crucially, QS governs EPS polymer gene clusters: LasR and RhlR directly regulate alginate biosynthesis genes, including algD, and modulate Pel and Psl biosynthesis operons, thereby linking intercellular signaling to biofilm polymer matrix composition.11
Ceftriaxone (CTX) is a third-generation cephalosporin antibiotic active against both Gram-positive and Gram-negative bacteria and is a drug of choice for treating pneumonia, meningitis, sepsis, and complicated urinary tract infections.12 The bactericidal and antibiofilm effects of CTX on P. aeruginosa have been reported in previous studies.13, 14 A previous study showed that sub-MICs of CTX inhibit biofilm formation through mechanisms involving QS.14 However, the structural basis for how CTX may directly interact with QS receptor proteins (LasR and RhlR) and biofilm EPS polymer biosynthetic enzymes at the molecular level has not been examined. Molecular docking is a validated computational approach that predicts the binding mode and affinity of a ligand within a target protein’s active site, enabling the rationalization of observed biological effects.15 Given the polymer-centric nature of P. aeruginosa biofilm resistance, elucidating the molecular interactions between CTX and biofilm-associated target proteins has direct relevance to the development of novel antibiofilm strategies and polymer-targeted therapies.
Objectives
The present study aims to evaluate the effect of sub-MICs of CTX on P. aeruginosa biofilm formation on abiotic surfaces and adhesion to mouse bladder epithelial cells; to elucidate the molecular mechanism of biofilm inhibition by assessing the effect of sub-MIC concentrations of CTX on QS regulatory genes (lasR and rhlR); and to perform docking analysis to investigate the binding interactions of CTX with the LasR and RhlR QS receptor proteins and biofilm EPS polymer biosynthetic enzymes (AlgD and PelB), thereby providing a structural basis for the observed antibiofilm activities.
Materials and methods
Bacterial isolation and identification
Midstream urine specimens were collected from 260 inpatients at Baghdad Teaching Hospital, Baghdad, Iraq, who had UTIs. The samples were collected in sterile containers. Patients were not allowed to receive antibiotic treatment within 3 days prior to sample collection. All patients signed the consent form before sample collection. The samples were inoculated onto MacConkey agar (HiMedia, Mumbai, India) and Cetrimide agar (HiMedia, Mumbai, India) plates and incubated at 37°C for 18 h. Biochemical tests (oxidase and catalase) and Gram staining were performed. The VITEK 2 system (bioMérieux, Marcy-l’Étoile, France) with the ID-GNB card was used for identification.16 Purified isolates were stored at −20°C in nutrient broth supplemented with 20% glycerol. The authors followed good laboratory practices when handling the biohazardous materials used in the current study.17
Antimicrobial susceptibility testing and minimum inhibitory concentration
Kirby–Bauer disk diffusion on Mueller–Hinton agar (MHA; HiMedia) using CTX disks (30 µg) was performed. Inhibition zone diameters were interpreted according to CLSI breakpoint guidelines.18 Minimum inhibitory concentrations (MICs) were determined using the broth microdilution method in Mueller–Hinton broth (MHB; HiMedia), with serial twofold dilutions of CTX (Panpharma, France) prepared from a 2 mg/mL stock solution.19
Biofilm formation assay
Biofilm formation was evaluated using the microtiter plate crystal violet assay on polystyrene 96-well plates. Briefly, 100 µL Tryptic Soy Broth (TSB, 0.25 g/L glucose, HiMedia, India) with 5 µL bacterial suspension (0.5 McFarland) was incubated at 37°C for 24 h. Wells were washed, fixed at 65°C for 60 min, stained with 0.4% crystal violet (15 min), washed, and destained with anhydrous ethanol. Absorbance at 590 nm (OD590) was measured with a microplate reader (BioTek 800 TS, Winooski, USA). Experiments were performed in triplicate, and biofilm classification was based on cut-off optical density values.14
Effect of CTX sub-MICs on biofilm polymer formation
The effect of CTX sub-MICs (1⁄2 MIC, 1⁄4 MIC, 1⁄8 MIC, 1⁄16 MIC, 1⁄32 MIC, and 1⁄64 MIC) on biofilm formation was assessed on polystyrene surfaces using the method described above. Three controls were included: TSB alone (negative), TSB with bacterial inoculum without antibiotic (positive biofilm control), and CTX serial dilutions in TSB without bacteria (background control). All experiments were performed in triplicate.
Effect of CTX sub-MICs on adhesion to mouse bladder epithelial cells
The effect of CTX sub-MICs (1⁄2 MIC, 1⁄4 MIC, 1⁄8 MIC, 1⁄16 MIC, and 1⁄32 MIC) on the adhesion of P. aeruginosa to mouse bladder epithelial cells (BECs) was evaluated. Mouse BECs were prepared in vitro by gently scraping the inner epithelial layer of the bladder obtained from healthy mice.20 Cells were maintained in Minimum Essential Medium (MEM; Sigma-Aldrich, St. Louis, USA) supplemented with 10% fetal bovine serum (Biowest, USA), 2 mM L-glutamine, and 1 mM sodium pyruvate. After incubation, non-adherent bacteria were removed by washing, and viable adherent bacteria were counted by serial tenfold dilution and plate counting on nutrient agar. Smears stained with Leishman stain were examined microscopically. Bacteria treated with phosphate-buffered saline (PBS; 0.1 M, pH 7.2) instead of CTX served as untreated controls.
Gene expression: qRT-PCR
The effects of sub-MICs of CTX (1⁄2 MIC, 1⁄4 MIC, 1⁄8 MIC, 1⁄16 MIC, and 1⁄32 MIC) on the expression of QS regulatory genes (rhlR and lasR) were evaluated using the isolate producing the highest level of biofilm formation. Total RNA was extracted with TRIzol reagent (Thermo Fisher Scientific, Waltham, USA). cDNA was synthesized with the RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific). qRT-PCR was performed on a Rotor-Gene Q thermocycler (Qiagen, Hilden, Germany) using TOPreal qPCR ×2PreMIX SYBR Green (Enzynomics, Daejeon, South Korea). Gene expression was normalized to the housekeeping gene rpoD using the 2^−ΔΔCT method.15, 21 Data are expressed as fold change relative to the PBS-treated control.
Molecular docking of ceftriaxone with biofilm-associated target proteins
In silico molecular docking was performed to investigate the binding interactions of CTX with QS receptor proteins (LasR and RhlR) and key biofilm EPS polymer biosynthetic enzymes (AlgD and PelB).
Ligand preparation: The three-dimensional structure of ceftriaxone (PubChem CID: 5479530; molecular formula: C₁₈H₁₈N₈O₇S₃; molecular weight: 554.58 g/mol) was retrieved from the PubChem database. The structure was converted to PDBQT format using Open Babel v. 3.1. Energy minimization was performed using the MMFF94 force field within Avogadro v. 1.2 prior to docking.
Target protein preparation: Crystal structures were downloaded from the Protein Data Bank (PDB): LasR ligand-binding domain in complex with N-(3-oxododecanoyl)-L-homoserine lactone (PDB: 2UV0, resolution: 1.9 Å), RhlR ligand-binding domain (PDB: 3T5K, resolution: 1.6 Å), AlgD GDP-mannose dehydrogenase (PDB: 1MV8), and PelB from the Pel biosynthetic operon (PDB: 5TCB). Water molecules and co-crystallized ligands were removed using PyMOL v. 2.5 (Schrödinger, New York, USA). Polar hydrogen atoms were added, and Gasteiger partial charges were assigned using AutoDockTools v. 1.5.7 (Scripps Research Institute, La Jolla, USA).
Docking simulations: Molecular docking was performed using AutoDock Vina v. 1.2 (Scripps Research Institute).22 The docking grid box was centered on the co-crystallized ligand binding site for each target (LasR: center x = 15.5, y = 18.3, z = 22.7; RhlR: center x = −4.2, y = 12.8, z = 5.4; AlgD: center x = 8.1, y = 22.5, z = −3.9; PelB: center x = 5.2, y = −8.7, z = 14.2), with box dimensions of 25 × 25 × 25 Å and an exhaustiveness parameter of 8. The top-ranked binding pose (lowest binding energy) was selected for analysis. Protein–ligand interactions were visualized using BIOVIA Discovery Studio Visualizer v. 21.1 (Dassault Systèmes, Vélizy-Villacoublay, France) and LigPlot+ v. 2.2 (EMBL-EBI, Hinxton, UK). Binding interactions (hydrogen bonds and hydrophobic contacts) were identified, and the interacting residues were recorded.22
Validation: The docking protocol was validated by re-docking the co-crystallized native ligand of each target protein into its respective binding site. Acceptable re-docking was confirmed by a root-mean-square deviation (RMSD) of ≤2.0 Å between the re-docked and crystallographic poses, consistent with standard validation criteria.
Statistical analyses
Origin v. 8.4 (OriginLab, Northampton, USA) was used to prepare graphs and perform data analysis. Data are presented as means ± standard error. Student’s t-test and one-way ANOVA were used to evaluate group differences. Correlation was assessed by Pearson’s correlation coefficient (r). A p < 0.05 was considered statistically significant.
Results
Bacterial isolates and CTX susceptibility
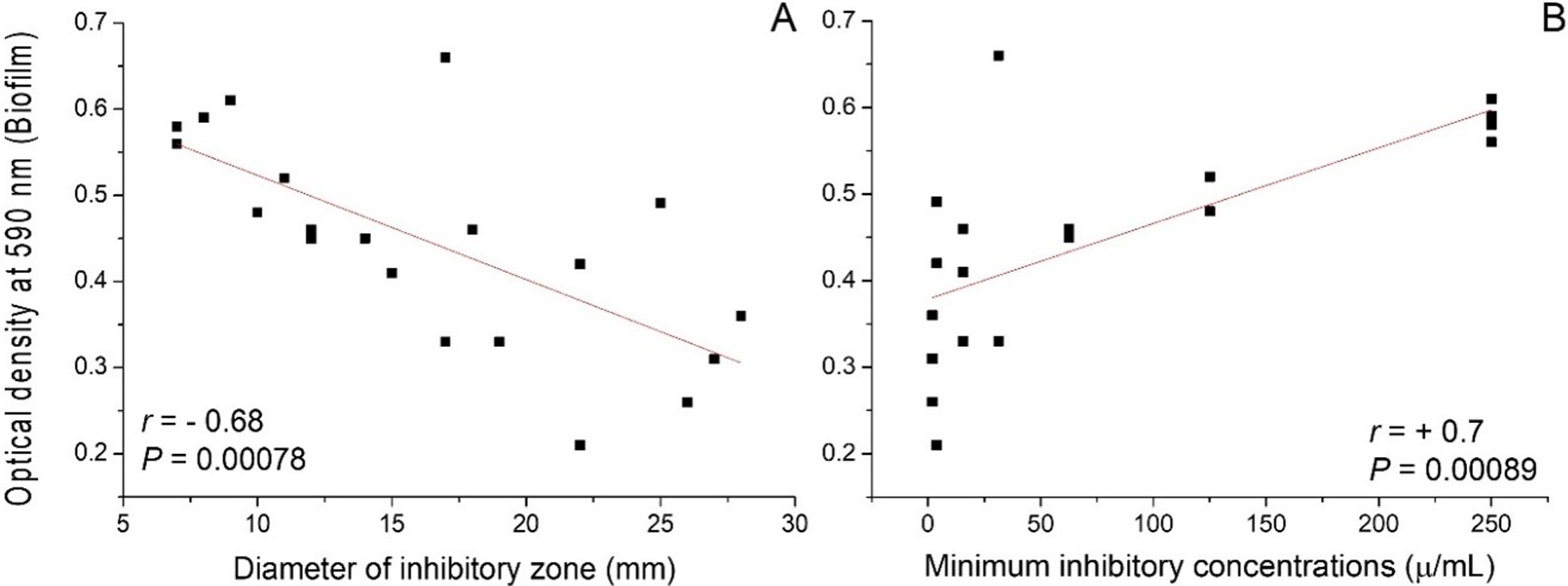
Twenty P. aeruginosa isolates were obtained from 260 midstream urine samples (incidence: 7.66%). The Kirby–Bauer disk diffusion method showed inhibitory zone diameters ranging from 7 ± 0.78 mm (Pa15) to 28 ± 4.8 mm (Pa7). MIC values ranged from 1.95 µg/mL (Pa2, Pa7, Pa17) to 250 µg/mL (Pa1, Pa5, Pa10, Pa15), confirming an inverse relationship between inhibition zone diameter and MIC (Table 1).
Biofilm formation
All 20 isolates produced biofilm to varying extents, with OD590 values ranging from 0.21 ± 0.17 to 0.66 ± 0.10. Five isolates (Pa1, Pa4, Pa5, Pa10, Pa15) were strong biofilm formers (OD590 > 0.50). Eight isolates were moderate biofilm formers, and four were weak biofilm formers. A significant negative correlation was observed between CTX inhibition zone diameter and biofilm formation (r = −0.68, p < 0.001), and a significant positive correlation was found between MIC values and biofilm biomass (r = +0.70, p < 0.001) (Figure 1). These findings indicate that strong biofilm formation is associated with reduced susceptibility to CTX, consistent with the protective role of EPS polymer matrices (alginate, Pel, Psl) in shielding bacterial cells from antibiotic penetration.
Effect of CTX sub-MICs on biofilm formation
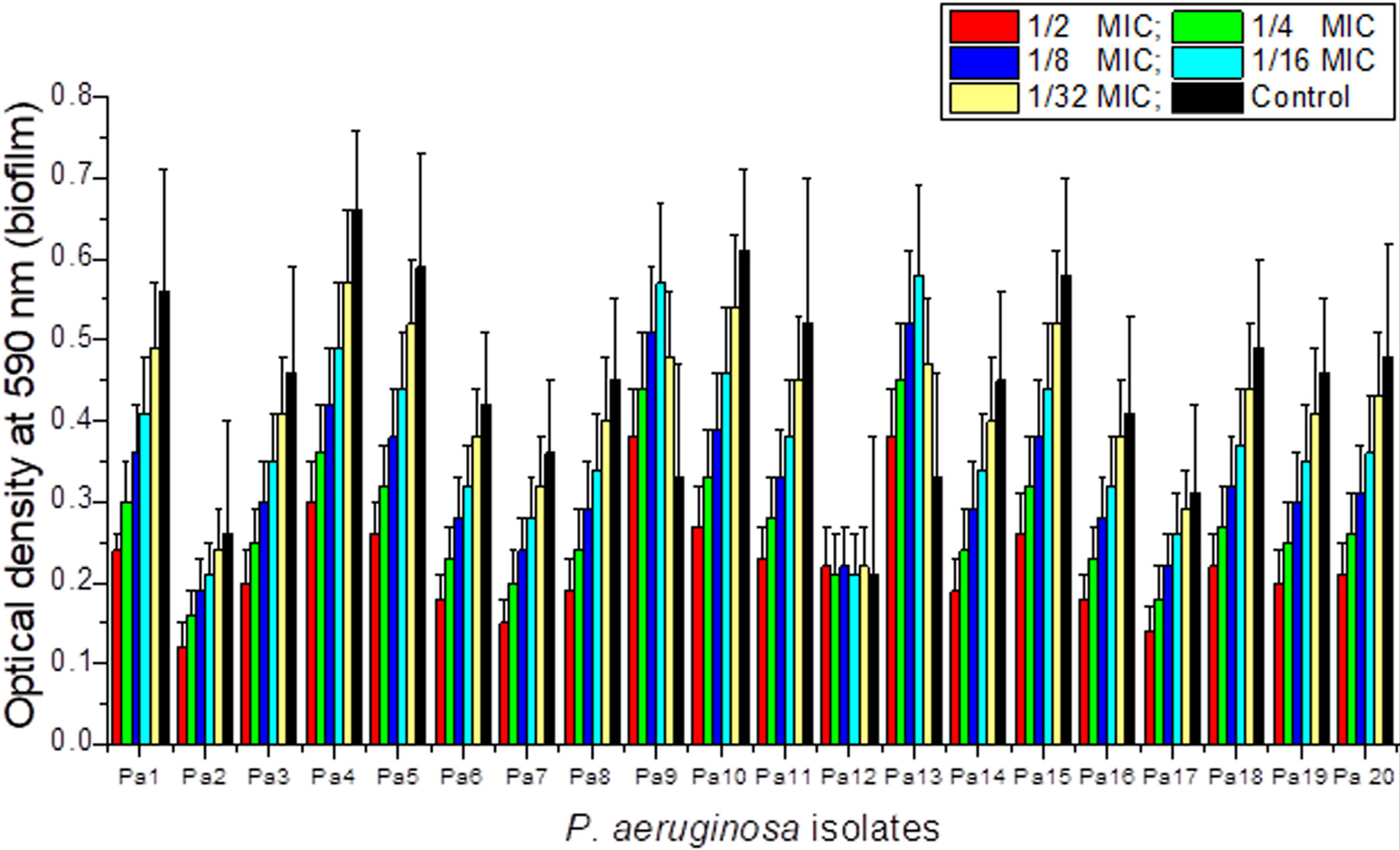
CTX sub-MICs (1⁄2, 1⁄4, 1⁄8, 1⁄16, 1⁄32 MIC) produced heterogeneous effects on biofilm formation across the 20 isolates. The majority of isolates (Pa1–Pa8, Pa10–Pa11, Pa14–Pa20) exhibited a concentration-dependent reduction in biofilm biomass. Two isolates (Pa9 and Pa13) showed a paradoxical, transient increase in biofilm formation at intermediate sub-MIC concentrations (1⁄8–1⁄16 MIC), suggesting a stress-induced adaptive biofilm response. One isolate (Pa12) showed no significant change across all CTX sub-MICs (Figure 2).
This paradoxical effect in a subset of strains likely reflects the upregulation of EPS polymer biosynthesis, particularly alginate overproduction, as a stress-response mechanism to a sublethal antibiotic challenge, a phenomenon associated with mucA mutation and sigma factor AlgT/U activation.23
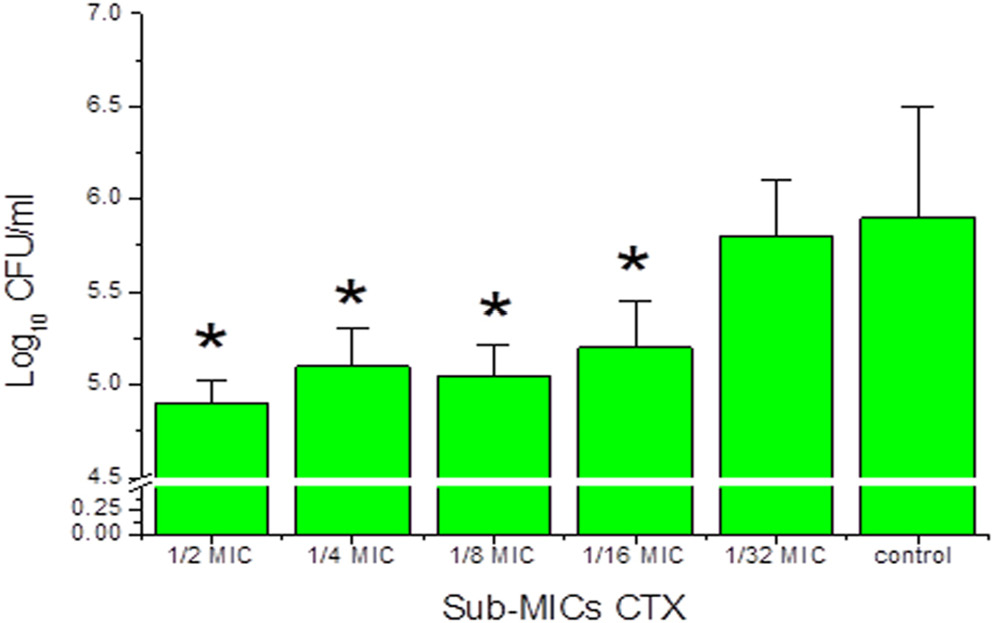
Effect of CTX sub-MICs on adhesion to mouse BECs
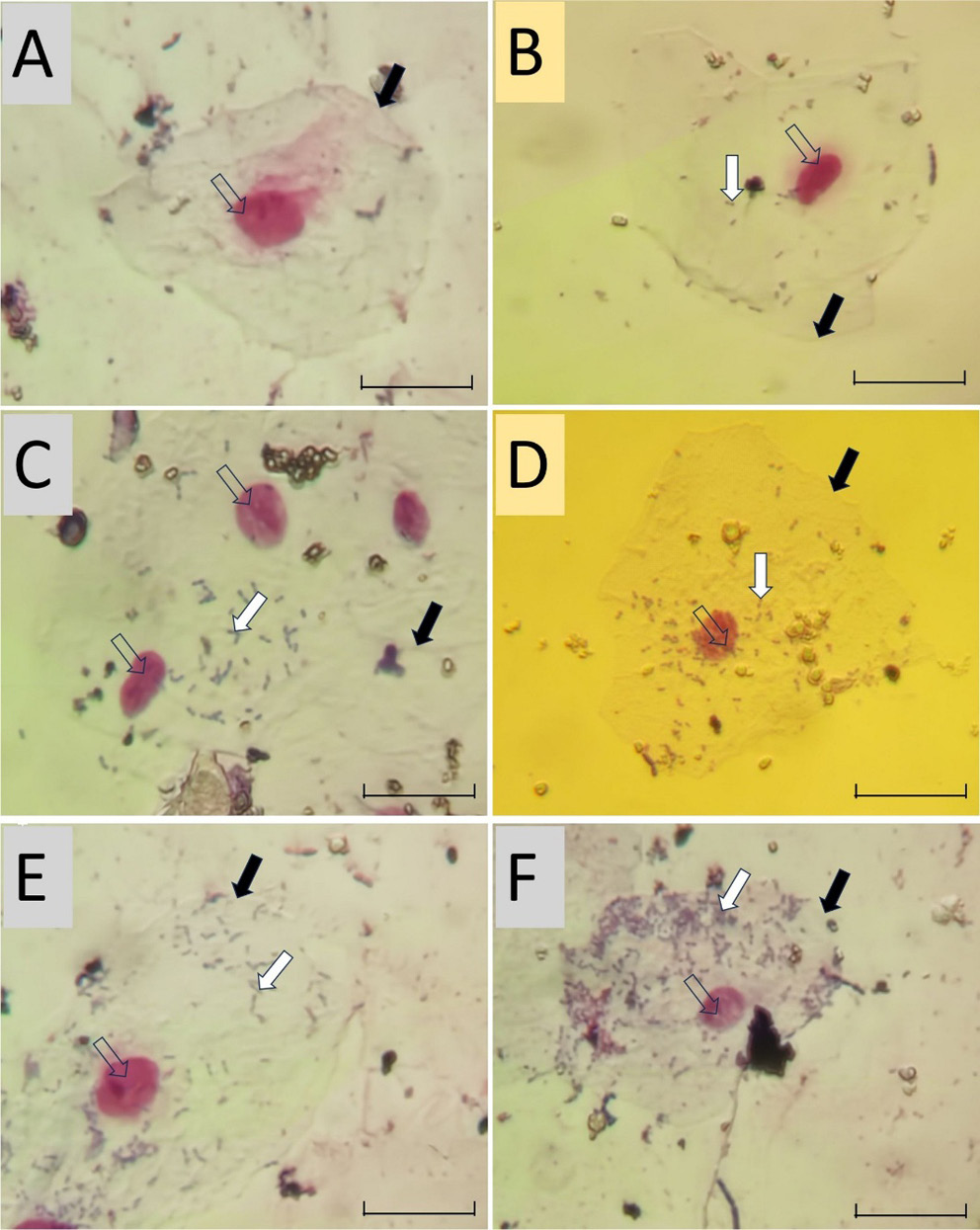
CTX sub-MICs reduced the number of viable Pa4 cells adherent to mouse BECs in a concentration-dependent manner. The greatest reduction was observed at 1⁄2 MIC. Significant decreases were observed at 1⁄2, 1⁄4, 1⁄8, and 1⁄16 MIC concentrations. No significant effect was observed at 1⁄32MIC (p > 0.05; fig. 3). Microscopic examination (Figure 4) confirmed a concentration-dependent reduction in bacterial adhesion to BECs. The reduction in surface adhesion at sub-inhibitory CTX concentrations may reflect diminished Psl polymer-mediated surface attachment, as Psl is a key mediator of initial surface colonization and its regulation falls under the QS network modulated by LasR and RhlR.
Effect of CTX sub-MICs on rhlR and LasR gene expression
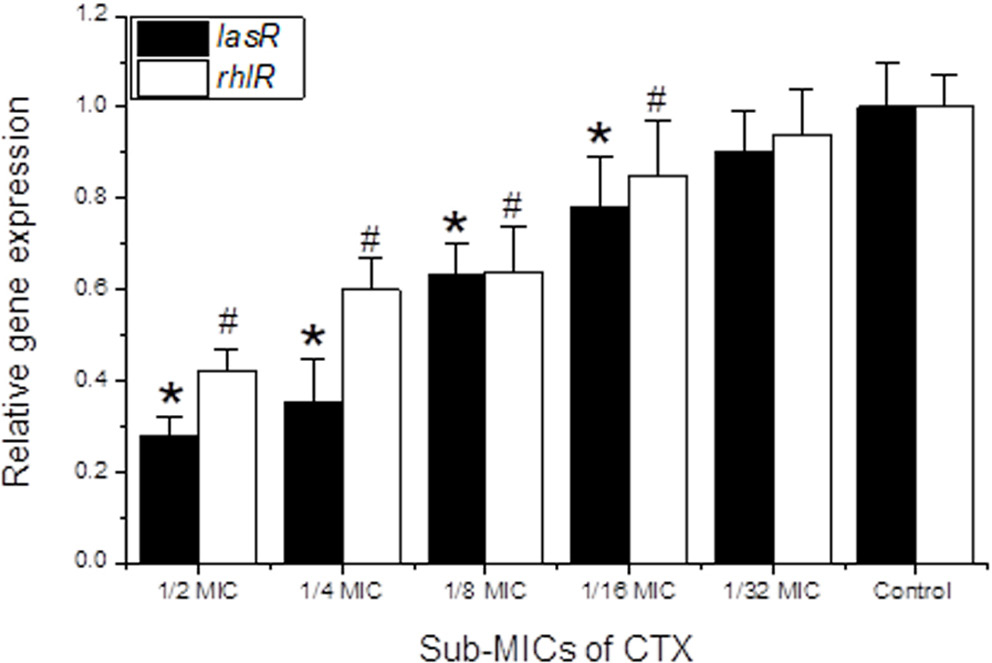
Sub-MICs of CTX significantly downregulated QS gene expression (rhlR and lasR) in Pa4 in a concentration-dependent manner (Figure 5). At ½ MIC, lasR and rhlR expression decreased to 0.28- and 0.42-fold, respectively. lasR exhibited greater sensitivity to CTX than rhlR at all sub-MIC concentrations. Since LasR directly governs the algD gene cluster and Pel/Psl biosynthesis operons, the observed downregulation of lasR by CTX sub-MICs provides a transcriptional mechanism for the concurrent reduction in biofilm EPS polymer matrix formation.
Molecular docking of ceftriaxone with biofilm-associated target proteins
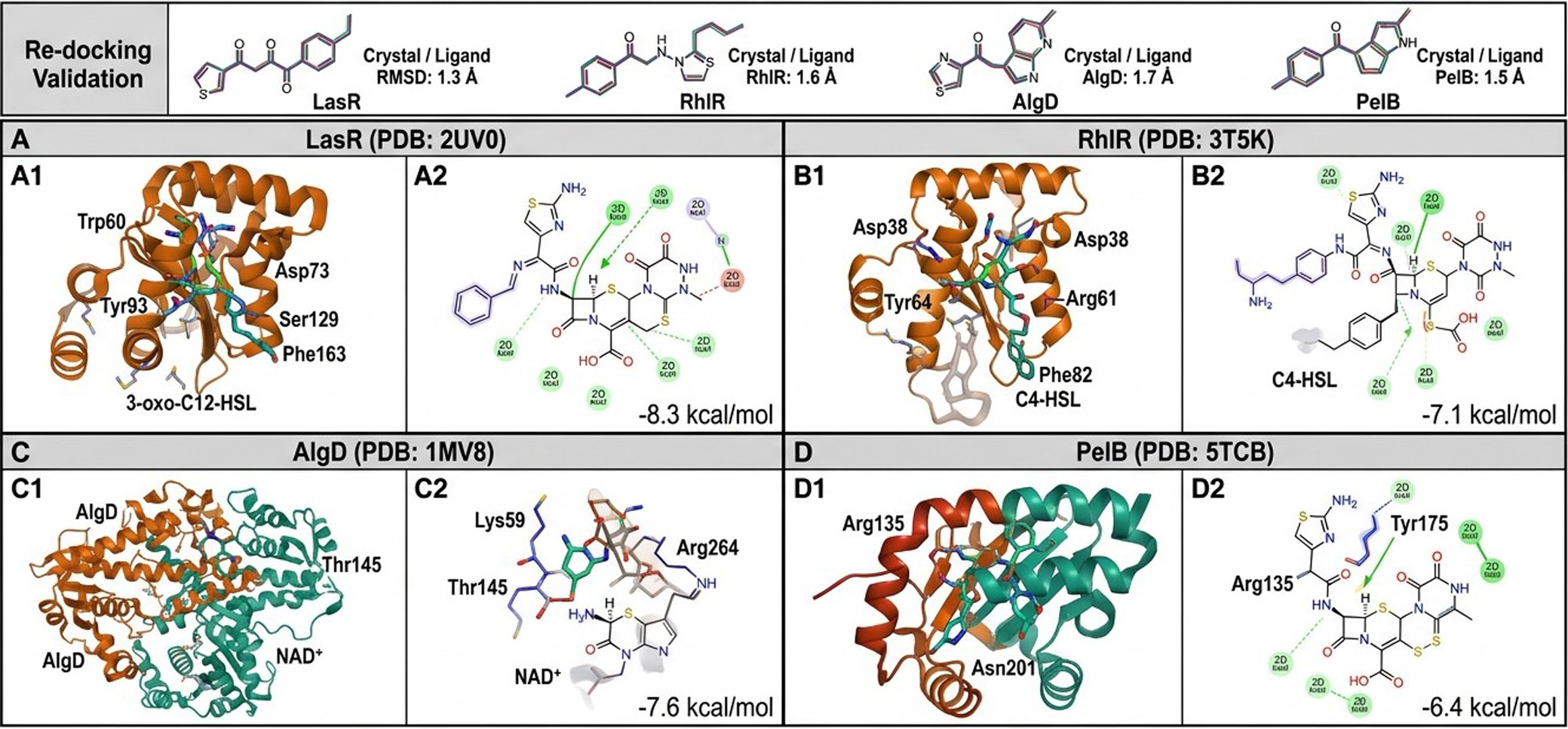
Re-docking validation: The re-docking RMSD values for all four target proteins were ≤1.8 Å (LasR: 1.3 Å; RhlR: 1.6 Å; AlgD: 1.7 Å; PelB: 1.5 Å), confirming the reliability of the docking protocol.
CTX–LasR interaction: Molecular docking of CTX into the LasR ligand-binding domain (PDB: 2UV0) yielded a binding energy of −8.3 kcal/mol, which is comparable to that of the native autoinducer N-(3-oxododecanoyl)-L-homoserine lactone (3-oxo-C12-HSL; −8.7 kcal/mol). CTX occupied the hydrophobic pocket of LasR, with hydrogen bond formation involving residues Trp60, Asp73, Tyr93, and Ser129 through its aminothiazole ring and oxime functional group. Hydrophobic contacts were established with Ala127, Tyr134, Leu125, and Phe163. The favorable docking energy and overlap with the autoinducer-binding site suggest that CTX may competitively displace the native QS signal from LasR, thereby inhibiting LasR-dependent transcriptional activation of virulence and EPS polymer biosynthesis genes (Table 2, Figure 6).
CTX–RhlR interaction: Docking of CTX into the RhlR ligand-binding domain (PDB: 3T5K) yielded a binding energy of −7.1 kcal/mol. Key hydrogen bonds were formed with Asp38, Tyr64, and Arg61, while hydrophobic contacts involved Ile40, Leu62, Phe82, and Val111. The lower binding energy compared with LasR is consistent with the experimentally observed greater sensitivity of lasR gene expression to CTX sub-MIC stress (Table 2, Figure 6).
CTX–AlgD interaction: AlgD (GDP-mannose dehydrogenase; PDB: 1MV8) is the rate-limiting enzyme in alginate polymer biosynthesis. Docking of CTX against AlgD yielded a binding energy of −7.6 kcal/mol, with key interactions involving the NAD+-binding residues Lys59, Thr145, and Arg264 via hydrogen bonding. These interactions were concentrated at the coenzyme-binding cleft, suggesting that CTX may interfere with the enzymatic activity of AlgD and thereby reduce alginate EPS polymer production (Table 2, Figure 6).
CTX–PelB interaction: Docking against PelB (PDB: 5TCB), a key outer-membrane auxiliary protein involved in Pel polysaccharide secretion, yielded a binding energy of −6.4 kcal/mol, with hydrogen bonding to Arg135, Tyr175, and Asn201. This interaction may interfere with Pel polymer export (Table 2, Figure 6). The binding energies are summarized in Table 2.
Discussion
UTIs caused by P. aeruginosa are among the most challenging conditions for physicians to manage owing to its extensive antibiotic resistance.24, 25 Biofilm formation is a central virulence mechanism, and the EPS polymer matrix comprising alginate, Pel, and Psl plays a decisive role in antibiotic tolerance and persistent infection.23, 26
The present study demonstrates that sub-MIC concentrations of CTX modulate P. aeruginosa biofilm formation and epithelial cell adhesion through QS gene downregulation and provides novel molecular docking evidence for direct CTX interactions with QS receptor proteins and biofilm EPS polymer biosynthetic enzymes.
In the present study, mouse BECs were used as a biotic model to study bacterial adhesion rather than human BECs because both cell types may yield comparable data on the adhesion of P. aeruginosa to biotic surfaces. The mouse BEC model was used because these cells are available in our laboratory, are easy to handle, and offer ethical advantages for preliminary adhesion studies.
All P. aeruginosa isolates from UTI urine samples formed biofilms at varying levels. The significant positive correlation between MIC and biofilm biomass (r = +0.70, p < 0.001) confirms that biofilm formation contributes to antibiotic tolerance. This correlation is mechanistically explained by the biofilm EPS polymer matrix: alginate and Pel polysaccharides create a physical diffusion barrier that reduces effective intra-biofilm CTX concentrations, while Psl reinforces surface adhesion. Isolates forming stronger biofilms thus require higher antibiotic concentrations for inhibition.27
Crystal violet staining is the preferred method for measuring biofilm biomass; it has been extensively validated in P. aeruginosa biofilm research and is used in several studies.13, 14, 18 This method measures the total biofilm biomass (including both bacterial cells and the biofilm extracellular matrix) and provides a statistically validated measure of treatment-induced suppression of biofilm formation.
Although direct quantification of EPS polymers would further support these data, we believe that combining biofilm biomass measurements with qRT-PCR gene expression analysis to determine how CTX affects biofilm formation yields valuable mechanistic insights into CTX-mediated biofilm modulation.
Sub-MICs of CTX reduced biofilm formation in the majority (17/20) of isolates in a concentration-dependent manner. Not only did CTX reduce biofilm formation, but previous studies have also shown that CTX possesses antibiofilm activity and that different β-lactam antibiotics can reduce biofilm formation by P. aeruginosa.
In the present study, the rhlR and lasR genes were chosen because they represent the 2 main quorum-sensing systems of P. aeruginosa. The 2 systems work together hierarchically to regulate biofilm formation and exopolysaccharide (EPS) production. The lasR gene is the master regulator of QS and activates the rhlR system, which in turn regulates the algD, pel, and psl operons. Therefore, these 2 genes are critical for understanding QS inhibition by CTX.28
At the molecular level, the concurrent downregulation of lasR and rhlR by CTX sub-MICs provides a transcriptional explanation for reduced EPS polymer production: LasR directly activates the algD biosynthetic gene cluster (required for alginate polymer synthesis) and the pel/psl operons responsible for Pel and Psl polysaccharide production.29 Downregulation of LasR therefore simultaneously reduces alginate viscosity, Pel-mediated cohesion, and Psl-mediated surface attachment, collectively dismantling the biofilm polymer scaffold.10, 11
Paradoxically, two isolates (Pa9 and Pa13) showed a transient increase in biofilm formation at intermediate sub-MIC concentrations. This stress-induced response is consistent with the well-characterized “alarm” mechanism in P. aeruginosa, whereby sublethal antibiotic stress triggers the AlgT/U sigma factor, leading to mucoid conversion and alginate overproduction as a protective response.
Importantly, the mucA gene encodes a negative regulator (anti-sigma factor) of AlgT/U, thereby suppressing alginate overproduction in P. aeruginosa. Under antibiotic stress, mucA is frequently mutated, resulting in constitutive overproduction of alginate and enhanced biofilm tolerance.23, 30
These findings underscore the risk of using subtherapeutic antibiotic concentrations, as they may drive selection for mucoid, hyper-biofilm-forming phenotypes and promote the emergence of new antibiotic-resistant P. aeruginosa isolates, making the treatment of bacterial infections a major challenge for physicians.
The present study demonstrated that sub-MICs of CTX downregulated the expression of lasR and rhlR in a concentration-dependent manner, with lasR exhibiting greater sensitivity. This hierarchy aligns with the upstream regulatory position of the Las system: LasR activates rhlR expression, meaning that CTX-mediated interference with LasR signaling has compounding downstream effects across the entire QS network.
Our molecular docking data provide a structural mechanism for this preferential sensitivity: CTX binds within the LasR ligand-binding domain with a binding energy (−8.3 kcal/mol) close to that of the native autoinducer (−8.7 kcal/mol), suggesting competitive displacement of 3-oxo-C12-HSL from the LasR binding pocket. This would prevent LasR dimerization and DNA binding, thereby repressing LasR-dependent transcription of virulence and EPS polymer biosynthesis genes.31
The molecular docking interaction of CTX with AlgD (binding energy: −7.6 kcal/mol) at the NAD+-binding site is particularly significant. AlgD catalyzes the irreversible oxidation of GDP-mannose to GDP-mannuronic acid, the committed step in alginate polymer biosynthesis. Inhibition of AlgD would reduce the pool of alginate monomers, directly impairing alginate EPS polymer production. Similarly, CTX interaction with PelB (−6.4 kcal/mol), an auxiliary protein involved in Pel polysaccharide secretion, suggests an additional mechanism by which CTX may disrupt Pel polymer assembly at the bacterial outer membrane.32, 33
These computational findings collectively suggest that CTX exerts antibiofilm effects through a multi-target mechanism: (i) QS receptor antagonism (LasR and RhlR), (ii) transcriptional downregulation of EPS polymer gene clusters, and (iii) potential direct enzymatic interference with alginate and Pel biosynthetic enzymes.
A previous study by Talib and Ghafil demonstrated the effects of CTX sub-MICs on P. aeruginosa biofilm formation and adhesion to human oral mucosal epithelial cells,14 while Naga et al. reported QS gene modulation by a ceftriaxone–nickel complex.15 The present study extends these findings by focusing on UTI-derived isolates, mouse BEC adhesion, and, critically, by providing the first molecular docking analysis of CTX binding to P. aeruginosa QS receptors and EPS polymer biosynthetic enzymes, offering a mechanistic structural framework for the observed biological activities. These docking insights align with emerging polymer-focused antibiofilm strategies that target EPS biosynthesis as a therapeutic vulnerability.34
The molecular docking analysis in the present study showed favorable binding energies and suitable binding-site localization for CTX interactions with LasR, RhlR, AlgD, and PelB. However, the current predictions remain computational and cannot be confirmed without direct biophysical evidence. In contrast to QS gene downregulation (validated by qRT-PCR), the proposed CTX–protein interactions remain mechanistic hypotheses that require validation using surface plasmon resonance (SPR), isothermal titration calorimetry (ITC), or enzyme inhibition assays. Therefore, QS inhibition via lasR/rhlR transcriptional suppression represents the primary validated mechanism of antibiofilm activity identified in this study, whereas targeting of EPS biosynthetic enzymes (AlgD and PelB) should be considered a plausible secondary mechanism pending experimental validation.
Limitations
This study has several limitations. First, only a single high-biofilm-forming isolate (Pa4) was used for QS gene expression and adhesion experiments, as the study aimed to characterize the effects of multiple sub-MIC concentrations rather than a single concentration. However, several previous studies used a single isolate to examine the effects of antibiotics on biofilm formation, and other studies investigated QS gene expression in a single bacterial isolate (P. aeruginosa and Vibrio harveyi).13, 18, 35, 36
Second, the molecular docking analysis is computational in nature and requires experimental validation by surface plasmon resonance, isothermal titration calorimetry, or co-crystallography to confirm direct CTX binding to LasR, RhlR, AlgD, and PelB proteins. Third, all experiments were conducted in vitro, and the relevance of sub-MIC CTX effects on biofilm EPS polymer dynamics in vivo requires further investigation in animal infection models. In our laboratory, we plan to extend the findings of the current study using a murine animal model. The project has been prepared and will commence upon receipt of final approval.
Finally, this study did not directly quantify EPS polymer components (alginate, Pel, and Psl) by chemical analysis, which would strengthen the correlation between QS gene downregulation and EPS polymer reduction. In future studies, we suggest evaluating existing QS inhibitors (e.g., furanones and salicylate analogs) and biofilm disruptors (e.g., DNase and chelating agents); the potential of CTX–QS inhibitor combinations remains unexplored. This approach would have high clinical relevance for the treatment of recalcitrant UTIs.
Conclusions
The present study shows that P. aeruginosa isolates from urine samples form variable levels of biofilm. Sub-inhibitory concentrations of CTX exerted heterogeneous effects on biofilm formation but reduced biofilm formation in most (17/20) isolates and decreased P. aeruginosa (Pa4) adhesion to mouse BECs.
The sub-inhibitory concentrations of CTX downregulated the QS regulatory genes lasR and rhlR, with lasR being more sensitive. Molecular docking analysis demonstrates that CTX binds favorably to the ligand-binding pockets of the LasR and RhlR QS receptor proteins, competitively displacing native autoinducers and providing a structural basis for QS inhibition.
Additional docking interactions with AlgD and PelB suggest that CTX may directly interfere with alginate and Pel EPS polymer biosynthesis. These findings position CTX as a multitarget antibiofilm and anti-QS agent, with potential implications for polymer-focused therapeutic strategies targeting P. aeruginosa biofilm EPS matrix formation.
Data availability
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request. Molecular docking files (input/output) are available upon request.
Consent for publication
Not applicable. No personal data, images, or information that could identify individuals are included in this manuscript.
Ethical approval
This study was approved by the Ethics Committee of the University of Baghdad, Iraq (CSEC/1025/0122; October 12, 2025), since it was a retrospective analysis of routinely collected clinical data; individual patient consent was waived in accordance with national ethical guidelines and the Declaration of Helsinki.